1.流行の概要
2007/08シーズンの流行の始まりは例年より1カ月以上早く、1987年のサーベイランス開始以来最も早い流行の立ち上がりであった。患者発生数のピークは第5週目にみられたが、ピーク時の患者数は過去10シーズンで2000/01シーズンに次いで2番目に少なく、流行規模も例年より小さかった。全国のサーベイランスネットワークから 4,476株のウイルスが分離され、流行の主流はAH1亜型で全分離数の82%を占め、AH3亜型、B型はそれぞれ11%、7%であった。
A/H1N1ウイルスは、2002/03シーズンから2シーズンは流行がみられなかったが、その後4シーズン続けての流行となり、2007/08シーズンは流行の主流を占めた。流行株の大半はワクチン株A/Solomon Islands/3/2006と類似の抗原性であったが、抗原変異株であるA/Brisbane/59/2007類似株が増加する傾向がみられた。
2007年11月から欧州を中心に抗インフルエンザ薬オセルタミビル耐性H1N1株の流行が起こり、ノルウェーでは分離株の67%は耐性であった。この耐性株はノイラミニダーゼ(NA)蛋白にH275Yという特徴的なアミノ酸置換をもっていた。これら耐性株は今冬のワクチン株A/Brisbane/59/2007と抗原的に類似しており、また、別の抗インフルエンザ薬ザナミビルには感受性であった。2008年に入ってからは、オセルタミビル耐性株が世界中で分離されるようになり、南アフリカ、中米(ホンジュラス、ニカラグア)などではH1N1分離株のすべては耐性であり、オーストラリア(分離株の74%)、ロシア(分離株の45%)などでも耐性株が急速に増加してきている。現時点で世界のH1N1分離株の35%はオセルタミビル耐性となっている(http://www.who.int/csr/disease/influenza/oseltamivir_summary/en/index.html)。
一方、世界のオセルタミビル生産量の70%近くを使用しているわが国でも緊急サーベイランスを行った(IASR 29: 155-159, 2008)。最終的に解析した1,713株のH1N1株のうち、44株(2.6%)が耐性であり、諸外国より耐性株の出現頻度は顕著に少なかった。現時点でH3N2およびB型には耐性株は見つかっていないが、今冬の流行において、わが国も諸外国のように耐性株が増加するのか継続した監視が必要である。
例年、流行の主流を占めてきたA/H3N2ウイルスは、過去12シーズンでは最も分離株数が少なく、分離株全体に占める割合も最低であった。前シーズン後半よりワクチン株A/Hiroshima(広島)/52/2005から抗原性の変化した株が分離されたが、2007/08シーズンには、この抗原変異株の代表株A/Brisbane/10/2007に類似した株が大半を占めるようになった。
前シーズンに大きな流行となったB型株は、分離株全体に占める割合が最も低く、大半はシーズンの後半に分離された。そのほとんどは前シーズンに流行したVictoria系統とは異なり、山形系統に属する株であった。これら分離株の多くは、抗原解析および遺伝子解析の結果から今冬のワクチン株B/Florida/4/2006に類似したB/Sendai(仙台)/114/2007類似株であった。一方、諸外国ではVictoria系統株と山形系統株の分離比率は地域により異なったが、世界的にみると山形系統株が流行の主流であった。
2.ウイルス抗原解析
2007/08シーズンに全国の地方衛生研究所(地研)で分離されたウイルス株は、各地研において、国立感染症研究所(感染研)からシーズン前に配布された抗原解析用抗体キット[A/Solomon Islands/3/2006(H1N1)、A/Hiroshima (広島)/52/2005(H3N2)、B/Shanghai(上海)/361/2002(山形系統)、B/Malaysia/2506/2004(Victoria系統)]を用いた赤血球凝集抑制(HI)試験によって、型・亜型別同定および抗原解析が行われた。感染研ではこれらの成績をもとにして、HI価の違いの比率が反映されるように選択した分離株(分離総数の約5~10%に相当)および非流行期の分離株や大きな抗原変化を示す分離株について、A/H1N1ウイルスに対しては6~8種類、A/H3N2ウイルス8~9種類、B型ウイルス8~9種類のフェレット参照抗血清を用いて詳細な抗原解析を行った。
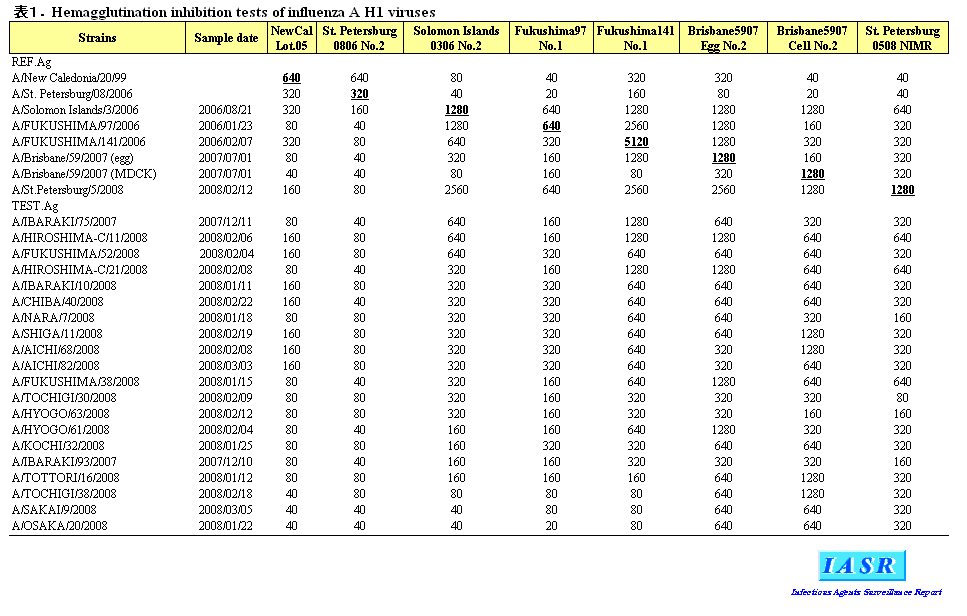
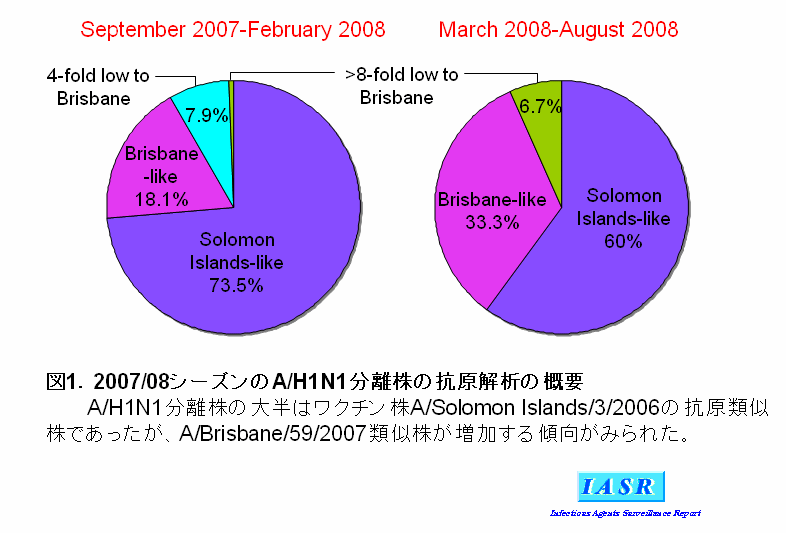
1)A/H1N1ウイルス:2007/08シーズンにはAH1亜型ウイルスは全国で3,646株分離された。感染研で解析した分離株の大半はワクチン株であるA/Solomon Islands/3/2006と抗原性が類似していたが、A/Brisbane/59/2007に代表される抗原変異株も多くみられた(表1)。これら変異株の割合はシーズン後半(2008年3月以降)には33.3%に達し、今冬も増加する傾向が見られた(図1)。
諸外国においては、欧州を中心にカナダ、南アフリカ共和国などでA/H1N1ウイルスの大きな流行がみられた。分離株はシーズン前半にはA/Solomon Islands/3/2006類似株が大半を占めていたが、シーズン後半には変異株であるA/Brisbane/59/2007類似株が多数を占めるようになった。すなわち、世界的にも流行の主流はA/Solomon Islands/3/2006類似株からA/Brisbane/59/2007類似株へと移行する傾向がみられた。
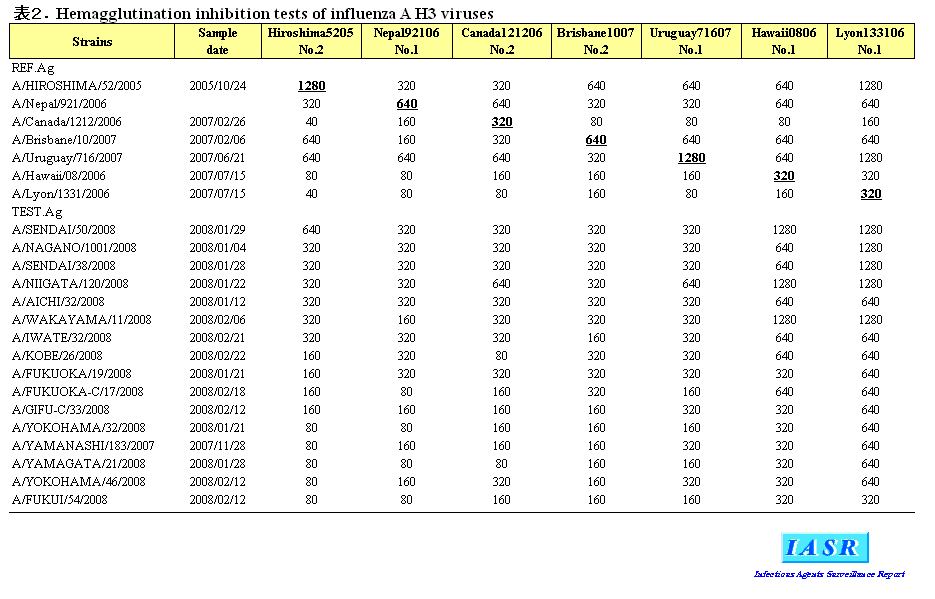
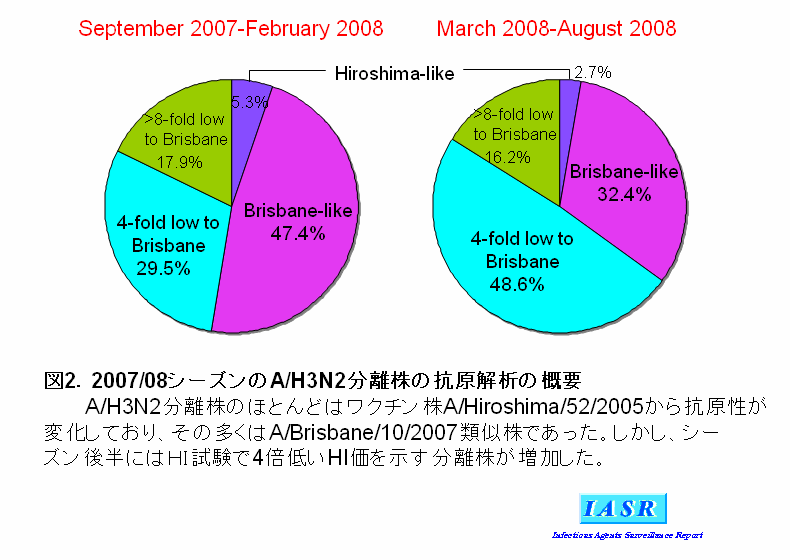
2)A/H3N2ウイルス:2007/08シーズンにはAH3亜型ウイルスは全国で504株分離された。感染研で解析した132株のほとんどは、ワクチン株であるA/Hiroshima(広島)/52/2005に対するフェレット感染血清とはHI試験で4倍以上の違いを示した(表2)。これら抗原変異株の半数はA/Brisbane/10/2007に対する感染血清によく反応することから、流行株の主流はA/Brisbane/10/2007類似株であることが示された(表2)。しかしながら、シーズン後半にはA/Brisbane/10/2007からHI試験で4倍低いHI価を示す分離株の割合が増え、A/Brisbane/10/2007からも抗原性が変化しつつあることが示唆された(図2)。
諸外国については、米国でA/H3N2亜型の大きな流行がみられたが、欧州やアジア諸国では大きな流行はなかった。2007/08シーズンの初めにはワクチン株のA/Wisconsin/67/2005類似株もみられたが、流行株のほとんどはA/Brisbane/10/2007やA/Uruguay/716/2007に代表される抗原変異株であった。
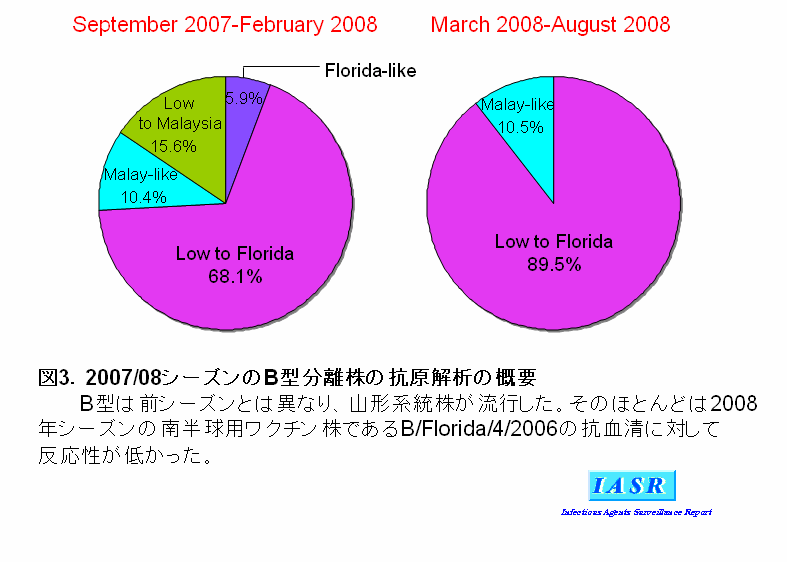
3)B型ウイルス:B型インフルエンザウイルスには、B/Yamagata(山形)/16/88に代表される山形系統とB/Victoria/2/87に代表されるVictoria系統がある。2007/08シーズンは3シーズンぶりに山形系統株がB型ウイルスの主流となり、B型ウイルス分離株総数(306株)の77%を占めた。感染研で解析した分離株のほとんどは、2008年の南半球用ワクチン株であるB/Brisbane/3/2007やB/Florida/4/2006に対するフェレット感染血清には反応性が低かったものの、抗原的にも遺伝系統的にもこれらの類似株であるB/Sendai(仙台)/114/2007に対する抗血清とよく反応することから、B/Florida/4/2006類似株であると判断された(表3、図3)。一方、少数ながら分離されたVictoria系統株の大半はワクチン株のB/Malaysia/2506/2004類似株であるB/Hiroshima(広島)/1/2005に対するフェレット感染血清とよく反応し、ワクチン類似株と判断された(表3、図3)。
諸外国においては、オランダ、ルクセンブルク、韓国やブラジルなど、一部の国を除くとB型ウイルスの流行はさほど大きくなかった。山形系統株とVictoria系統株の混合流行がみられ、両者の割合は地域によって異なったが、世界的には山形系統が主流を占めた。しかし、2008年の夏以降、オーストラリア、ニュージーランドや香港など、一部の国ではVictoria系統株が増加する傾向もみられた。
山形系統分離株の多くはB/Florida/4/2006類似株であった。一方、Victoria系統分離株の大半はワクチン株B/Malaysia/2506/2004の抗原類似株であったが、変異株も増加しつつある傾向がみられた。
3.ウイルス遺伝子解析
1)A/H1N1ウイルス:HA遺伝子の系統樹解析では(図4 pdf)、2007/08シーズンのすべてのH1N1株はT82K、Y94H、R145K、R208K、T266Nのアミノ酸置換をもつ一群(クレード2)に属した。クレード2内はさらに3つのサブクレードを形成し、K73R、R145Kのアミノ酸置換をもち代表株としてA/Solomon Islands/3/2006を含む群(サブクレード2A)、D35N、R188K、E273Kのアミノ酸置換を持ち代表株としてA/Brisbane/59/2007を含む群(サブクレード2B)および、S36N、R145K、R188M、A189T、T193Kのアミノ酸置換をもち代表株としてA/Hong Kong(香港)/2652/2006を含む群(サブクレード2C)にわかれた。今シーズンの国内および周辺アジア諸国で分離されたH1N1株はサブクレード2Bまたはサブクレード2Cに属し、サブクレード2Aに属する株は見られなかった。一方、ヨーロッパ、北米、オーストラリアで分離された最近のH1N1株の大多数はサブクレード2Bに属すため、2Cに属する株のほとんどはアジア分離株であった。2007/08シーズンに世界各地で分離されている抗インフルエンザ薬オセルタミビル耐性株のほとんどすべてはサブクレード2Bに属した。NA遺伝子の系統樹は省略するが、NA遺伝子もHA遺伝子と同様で、2007/08シーズンの分離株はサブクレード2Bまたはサブクレード2Cに属していた。
2)A/H3N2ウイルス:2007/08シーズンのすべてのH3N2株は、G50E、K140Iのアミノ酸置換をもち代表株としてA/Brisbane/10/2007およびA/Uruguay/716/2007を含むBrisbane-10系統に属した(図5 pdf)。Brisbane-10系統内には、K173Nのアミノ酸置換を持ち代表株としてA/Saitama(埼玉)/27/2008を含む群(グループ1)と、K173Qのアミノ酸置換を持ち代表株としてA/Toyama(富山)/123/2008を含む群(グループ2)の2つの大きなグループがあり、最近のH3N2株の大多数がどちらかのグループに属した。その他、小さな2つのグループとしてK173Eアミノ酸置換を持つ群と、N171Kのアミノ酸置換を持つ群があり、少数の最近のH3N2株はこれらのグループに属した。NA遺伝子についてもHA遺伝子と同様の傾向が見られた(図は省略)。
3)B型ウイルス:B型ウイルスは前述したように山形系統とVictoria系統に大別される。2007/08シーズンのVictoria系統株のほとんどはK48E、K80R、K129N のアミノ酸置換を持ち代表株としてB/Malaysia/2506/2004を含む一群に属した(図6 pdf)。
一方、2007/08シーズンの山形系統株のすべては、V251Mをもつ一群に属した(図7 pdf)。この群はさらに、3つのサブグループを形成し、G229Sアミノ酸置換をもち代表株としてB/Florida/4/2006を含む群(サブグループ1)、P108Aのアミノ酸置換を持ち代表株としてB/Brisbane/3/2007を含む群(サブグループ2)および、N165Y、G229Dのアミノ酸置換をもち代表株としてB/Bangladesh/3333/2007を含む群(サブグループ3)に分かれ、最近の山形系統株はいずれかのグループに属した。両系統間のHA・NA遺伝子交雑ウイルスは、2007/08シーズンの分離株中には見られなかった。
本研究は「厚生労働省感染症発生動向調査に基づくインフルエンザサーベイランス」事業として全国76地研と感染研ウイルス第3部第1室(インフルエンザウイルス室)との共同研究として行われた。また、インフルエンザウイルスの遺伝子解析は「インフルエンザウイルス遺伝子の大量解析に関する事業」として独立行政法人製品評価技術基盤機構と感染研ウイルス第3部第1室との共同研究として行われた。本稿に掲載した成績は全解析成績の中から抜粋したものであり、残りの成績は既に感染症サーベイランスシステム(NESID)の病原体検出情報システムで各地研に還元された。また、本稿は上記研究事業の遂行にあたり、地方衛生研究所全国協議会と感染研との合意事項に基づく情報還元である。
国立感染症研究所ウイルス第三部第一室・WHOインフルエンザ協力センター
独立行政法人製品評価技術基盤機構
 今月の表紙へ戻る
今月の表紙へ戻る
 IASRのホームページに戻る
IASRのホームページに戻る Return to the IASR HomePage(English)
Return to the IASR HomePage(English)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}